Modeling and mapping species distributions
Richard Chandler
Feb 5, 2019
Source:vignettes/spp-dist.Rmd
spp-dist.RmdAbstract
A species’ distribution can be characterized by either occurrence
probability or population density, defined for all locations in some
spatial extent. Defining distribution in terms of these two parameters
%These definitions of species distribution avoids the ambiguity
surrounding the indices of occurrence or abundance produced by many
presence-only algorithms. The unmarked package contains
methods of fitting occurrence and abundance models, and can be used to
produce distribution maps with the help of R’s GIS
capabilities, as is demonstrated in this vignette. Unlike many other
tools for modeling species distributions, the models in
unmarked account for bias due to spatial and temporal
heterogeneity in detection probability. Furthermore,
unmarked includes models of population dynamics, allowing
one to map quantities such as local colonization or extinction
probability.
Mapping Occurrence Probability
In this example, we use the occu function to fit the
single-season occupancy model of MacKenzie et al.
(2002) to data on the European crossbill (Loxia
curvirostra) collected in 267
1-km
sample quadrats in Switzerland, 1999 (Schmid et
al. 2004). We then use the model to compute the expected
probability of occurrence at each pixel in a raster defining the Swiss
landscape.
First we load the crossbill data, which contains the
detection/non-detection data and covariates. The dataset actually
contains data from multiple years, but we are only going to analyze data
from the first year, 1999. A multi-year analysis of occupancy dynamics
could be accomplished using the colext function, and in
addition to mapping occurrence probability, it would be possible to also
map colonization and extinction probabilities. The following commands
format the data.
library(unmarked)
data(crossbill)
umf <- unmarkedFrameOccu(
y=as.matrix(crossbill[,c("det991", "det992", "det993")]),
siteCovs=crossbill[,c("ele", "forest")],
obsCovs=list(date=crossbill[,c("date991", "date992", "date993")]))
sc <- scale(siteCovs(umf))
siteCovs(umf) <- sc
head(umf)## Data frame representation of unmarkedFrame object.
## y.1 y.2 y.3 ele forest date.1 date.2 date.3
## 1 0 0 0 -1.15390803 -1.14710440 34 59 65
## 2 0 0 0 -1.15390803 -0.49668159 17 33 65
## 3 NA NA NA -0.21745379 -0.09920098 NA NA NA
## 4 0 0 0 -0.37352950 -0.93029679 29 59 65
## 5 0 0 0 -0.06137809 0.00920282 24 45 65
## 6 NA NA NA -0.99783232 -1.18323900 NA NA NA
## 7 0 0 0 -0.68568091 -1.03870060 26 54 74
## 8 0 0 0 -0.84175662 0.91256783 23 43 71
## 9 0 0 0 -0.99783232 -1.07483520 21 36 56
## 10 0 0 0 -0.99783232 -0.78575839 37 62 75Notice that the site covariates, elevation and forest, were
standardized using the scale function. Standardization
isn’t always necessary, but it can make it easier to find the maximum
likelihood estimates. When standardizing covariates and then making
predictions, it is important to retain the original sample mean and
standard deviation. The reason for this is explained below.
Fitting a model is now straight-forward. In many cases, we would fit several models corresponding to competing hypotheses, but for simplicity, we stick with this single model.
## Warning: 22 sites have been discarded because of missing data.##
## Call:
## occu(formula = ~date ~ ele + I(ele^2) + forest, data = umf)
##
## Occupancy (logit-scale):
## Estimate SE z P(>|z|)
## (Intercept) 0.464 0.541 0.857 0.39137
## ele 1.228 0.319 3.846 0.00012
## I(ele^2) -1.333 0.417 -3.194 0.00140
## forest 0.654 0.274 2.385 0.01706
##
## Detection (logit-scale):
## Estimate SE z P(>|z|)
## (Intercept) -2.2356 0.52283 -4.28 0.000019
## date 0.0254 0.00752 3.39 0.000711
##
## AIC: 455.3892
## Number of sites: 245
## ID of sites removed due to NA: 3 6 38 63 71 106 116 118 125 126 152 163 168 178 181 197 199 212 218 220 238 257Now that we have our fitted model, we can use it to predict
occurrence probability at each pixel in the Swiss landscape. The
Switzerland dataset contains country-wide data. There are
many ways to display it—here is an example of mapping elevation using
the levelplot function in the lattice package
(Sarkar 2008).
library(lattice)
data(Switzerland)
print(levelplot(elevation ~ x + y, Switzerland, aspect="iso",
xlab="Easting (m)", ylab="Northing (m)",
col.regions=terrain.colors(100)))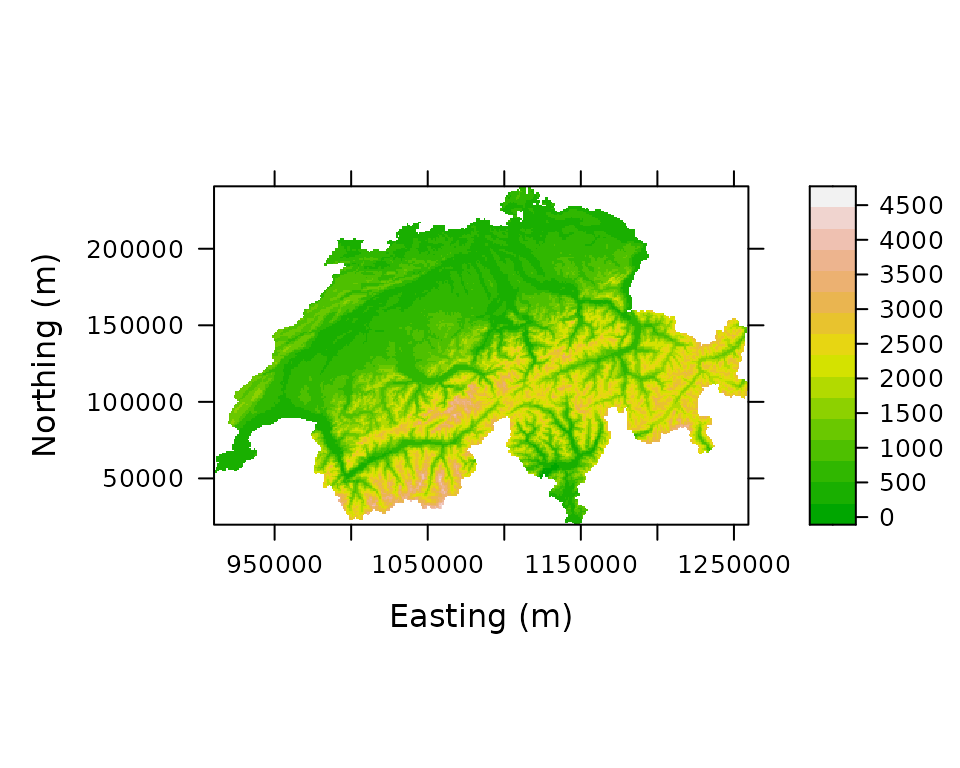
Figure 1. Elevation in Switzerland
The raster package (Jacob van
Etten 2012) provides another alternative. Here we create two
raster objects and specify the coordinate system.
elevation <- rasterFromXYZ(Switzerland[,c("x","y","elevation")],
crs="+proj=somerc +lat_0=46.95240555555556 +lon_0=7.439583333333333 +k_0=1 +x_0=600000 +y_0=200000 +ellps=bessel +towgs84=674.374,15.056,405.346,0,0,0,0 +units=m +no_defs")
forest <- rasterFromXYZ(Switzerland[,c("x","y","forest")],
crs="+proj=somerc +lat_0=46.95240555555556 +lon_0=7.439583333333333 +k_0=1 +x_0=600000 +y_0=200000 +ellps=bessel +towgs84=674.374,15.056,405.346,0,0,0,0 +units=m +no_defs")Since we standardized the covariates during the model fitting
process, we need to transform the country-wide data using the same
values. Note, we don’t want to use the mean and SD of the rasters
themselves, we want to use the mean and SD of the original covariates
used to fit the models, which are stored as attributes of the
sc object. The following commands display the original
means and SDs and then transform the rasters and join them in a raster
“stack”.
attr(sc, "scaled:center")## ele forest
## 1189.32584 34.74532
attr(sc, "scaled:scale")## ele forest
## 640.71471 27.67431
ele.s <- (elevation-1189)/640
forest.s <- (forest-34.7)/27.7
ef <- stack(ele.s, forest.s)
names(ef) <- c("ele", "forest")
plot(ef, col=terrain.colors(100))
Figure 2. Elevation and forest cover, standardized
It is important to assign names that exactly match the
covariate names used to fit the model. This is required by the
predict function as demonstrated later. The
predict function is useful for computing
spatially-referenced model predictions, standard errors, and confidence
intervals, but it is computationally demanding when there are many
pixels in the raster. Thus, if measures of uncertainty are not required,
the following code can be used to quickly produce the species
distribution map shown in Fig. 3.
(beta <- coef(fm.occu, type="state"))## psi(Int) psi(ele) psi(I(ele^2)) psi(forest)
## 0.4638233 1.2276426 -1.3327186 0.6544976
logit.psi <- beta[1] + beta[2]*ele.s + beta[3]*ele.s^2 + beta[4]*forest.s
psi <- exp(logit.psi) / (1 + exp(logit.psi))
print(spplot(psi, col.regions=terrain.colors(100)))
Figure 3. A species distribution map for the European crossbill in Switzerland. The colors represent occurrence probability.
As of version 0.9-6, the predict method in
unmarked can make predictions using an object of class
RasterStack from the package. As mentioned previously, the
rasters must be named, perhaps by using the
names(someraster) <- somename method. The object
returned by predict is another raster stack with rasters
for the expected values of the parameter of interest, the standard
errors, and the upper and lower confidence intervals. The following
example is very slow because there are many of pixels in the raster. The
resulting map is shown in Fig. 4.
E.psi <- predict(fm.occu, type="state", newdata=ef)
plot(E.psi, axes=FALSE, col=terrain.colors(100))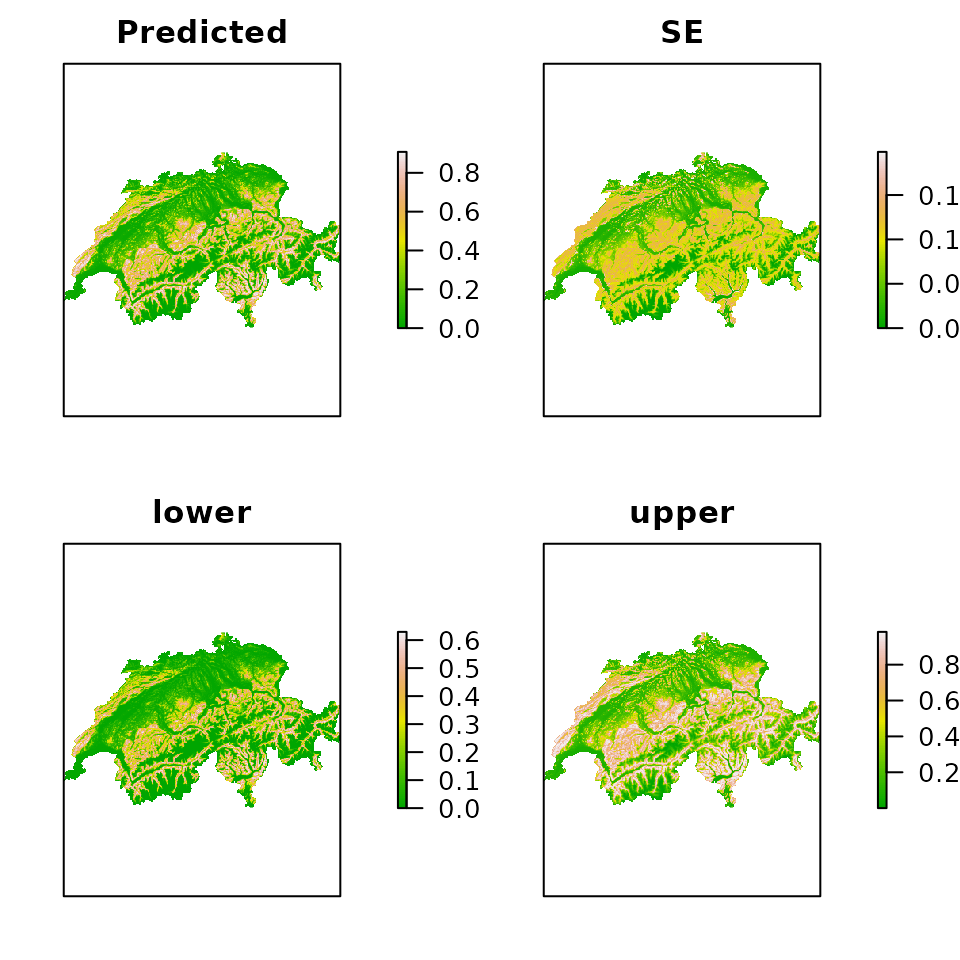
Figure 4. Expected occurrence probability along with standard errors and the limits of the asymptotic 95% confidence interval.
Users should be cautious when predicting from models that have
categorical predictor variables, i.e. factors. The package
does not have advanced methods for handling factors, and thus it is not
easy to automatically create dummy variables from them as can typically
be done using model.matrix. The safest option is to create
the dummy variables manually before fitting the models, and to use the
same variables as rasters for prediction.
A more important consideration when creating species distribution maps based upon occurrence probability is that of spatial scale. Occurrence probability will typically depend upon the area of the “site” in question. Thus, in our crossbill example, it would not be appropriate to use our model to predict occurrence probability for 10-km pixels since the surveys were done in 1-km quadrats. In some cases it might be possible to directly model the effect of site area on occurrence probability, in which case the effect could be accounted for in the predictions.
Mapping Population Density
Although distribution is typically described in terms of occurrence probability, which is always better than an index of occurrence probability, the best parameter for modeling species distribution is population density because density allows for inference about population size in any region of the species’ range. Furthermore, occurrence probability is simply the probability that abundance is greater than 0, so with density/abundance estimates, it is always possible to compute occurrence probability as a derived parameter.
In this example, we create a distribution map for the Island Scrub-Jay (Aphelocoma insularis), which is restricted to Santa Cruz Island, California. To do so, we fit the hierarchical distance sampling model of Royle et al. (2004), which allows for the estimation of abundance in each of the m pixels representing the island. The data were collected 307, 300-m radius point count (or “point transect”) surveyed during the Fall of 2008.
IMPORTANT: This analysis is for demonstration purposes only, and the estimates of population size should not be used for conservation or management purposes. Indeed, the Poisson assumption used here was found to be inadequate by Sillett et al. (2012) who conducted a rigorous analysis and reported reliable estimate of population size.
Although we are fitting a model of population density, the steps of the analysis closely mirror those shown in the previous section. First, we format the data and fit a model, then we format the island-wide covariates and make predictions. The three covariates thought to influence jay abundance are elevation, forest cover, and chapararral cover. We also include include the area of the survey plots in the analysis so that we can make predictions for regions of any area. Here is the code to format the data and fit the model.
data(issj)
covs <- scale(issj[,c("elevation", "forest", "chaparral")])
area <- pi*300^2 / 10000
jayumf <- unmarkedFrameDS(y=as.matrix(issj[,1:3]),
siteCovs=data.frame(covs, area),
dist.breaks=c(0,100,200,300),
unitsIn="m", survey="point")
head(jayumf)## Data frame representation of unmarkedFrame object.
## y.1 y.2 y.3 elevation forest chaparral area
## 1 0 0 2 -1.20607849 -0.3309618 -0.1218243 28.27433
## 2 0 0 0 -0.36132054 -0.4429552 0.8384709 28.27433
## 3 0 0 0 -0.45797193 -0.3738751 2.1319298 28.27433
## 4 0 0 0 -0.14196112 1.3908065 -0.2737078 28.27433
## 5 0 0 0 -0.72588125 -0.4869152 -1.1562240 28.27433
## 6 0 0 0 0.01704522 0.6706992 0.2989173 28.27433
## 7 0 0 0 -0.11866710 -0.2671150 1.0326122 28.27433
## 8 0 0 0 -0.76411740 -0.4921486 -0.9332983 28.27433
## 9 0 0 0 0.40065595 0.5440524 -1.0759952 28.27433
## 10 0 0 0 -0.24030211 -0.4921486 -1.1023299 28.27433
fm1 <- distsamp(~chaparral ~chaparral + elevation + offset(log(area)),
jayumf, keyfun="halfnorm", output="abund",
starts=c(-2.8,1,0,4.5,0))
fm1##
## Call:
## distsamp(formula = ~chaparral ~ chaparral + elevation + offset(log(area)),
## data = jayumf, keyfun = "halfnorm", output = "abund", starts = c(-2.8,
## 1, 0, 4.5, 0))
##
## Abundance (log-scale):
## Estimate SE z P(>|z|)
## (Intercept) -2.827 0.1609 -17.57 4.34e-69
## chaparral 0.957 0.1460 6.55 5.60e-11
## elevation -0.244 0.0932 -2.61 8.96e-03
##
## Detection (log-scale):
## Estimate SE z P(>|z|)
## (Intercept) 4.73 0.0845 55.96 0.000000
## chaparral -0.25 0.0744 -3.36 0.000773
##
## AIC: 964.6426
## Number of sites: 307
##
## Survey design: point-transect
## Detection function: halfnorm
## UnitsIn: m
## UnitsOut: haRemarks. 1) The distance data were binned into 3 distance classes. 2)
We used output="abund" even though, by specifying the
offset, we effectively modeled population density. As stated previously,
this allows us to make predictions of abundance for regions of arbitrary
size.
The next thing to do is to format the raster data. For details, see
the previous section-the process is the same, except that we need a
raster for "area", the size of each pixel in the raster
data. This is necessary because the survey plots were larger than the
pixels for which we want predictions of abundance.
data(cruz)
elev <- rasterFromXYZ(cruz[,c("x","y","elevation")],
crs="+proj=utm +zone=11 +ellps=GRS80 +datum=NAD83 +units=m +no_defs")
forest <- rasterFromXYZ(cruz[,c("x","y","forest")],
crs="+proj=utm +zone=11 +ellps=GRS80 +datum=NAD83 +units=m +no_defs")
chap <- rasterFromXYZ(cruz[,c("x","y","chaparral")],
crs="+proj=utm +zone=11 +ellps=GRS80 +datum=NAD83 +units=m +no_defs")
area.raster <- chap
values(area.raster) <- 300*300/10000 # area of a grid pixel
attr(covs, "scaled:center")## elevation forest chaparral
## 202.0023616 0.0673357 0.2703592
attr(covs, "scaled:scale")## elevation forest chaparral
## 124.8818069 0.1368199 0.2338295
elev.s <- (elev-202)/125
forest.s <- (forest-0.0673)/0.137
chap.s <- (chap-0.270)/0.234
habitat <- stack(elev.s, forest.s, chap.s, area.raster)
names(habitat) <- c("elevation", "forest", "chaparral", "area")Now, when we use predict, it will return the expected
number of jays in each pixel along with the standard errors and the 95%
confidence intervals. We could sum these up to obtain an estimate of
total population size. Sillett et al.
(2012) did this and used the parametric bootstrap to estimate the
variance of total population size.
E <- predict(fm1, type="state", newdata=habitat)
plot(E, axes=FALSE, col=terrain.colors(100))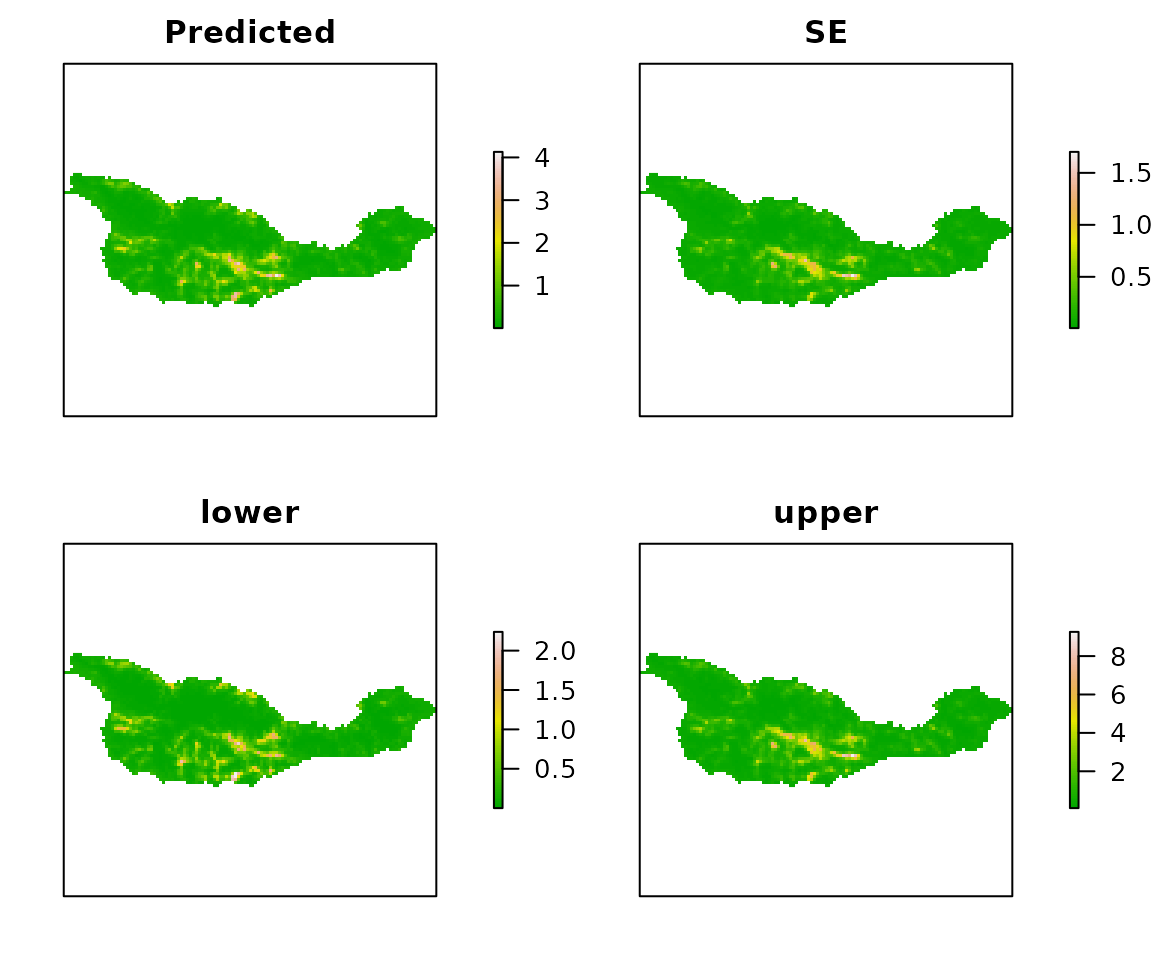
Figure 5. Expeted Island Scrub-Jay abundance, SEs, and 95% CIs.